Project 2: Understanding the Cellular Origins of Melanoma
PIs: Anand Ganesan, Arthur Lander; Key Personnel: Devon A. Lawson, John Lowengrub, Bruce Tromberg, Tatiana Krasieva
Melanoma, a tumor resistant to therapy in late stages, is curable by excision when caught early. Early melanomas can be difficult to distinguish from benign, pigmented “moles”, i.e. melanocytic nevi; this leads to unnecessary excision of many normal nevi while early melanomas are often missed. Nevi and melanomas share more than morphological features: Clinical and experimental data show that ~90% of nevi are initiated when melanocytes acquire an activating mutation in the BRAF oncogene, the same oncogenic mutation observed in >60% of melanomas. Yet nevi spontaneously stop growing. This is usually attributed to “oncogene-induced senescence,” but the fact that nevi readily re-grow after incomplete excision, or in response to UV-irradiation, and can sometimes evolve to melanoma, suggest nevi are not “senescent” but reversibly growth-arrested. Nevi also spontaneously regress, a process that appears to involve the immune system.
In preliminary work, we investigated nevus dynamics in a mouse model of inducible Braf activation, which mimics human nevus formation and also produces melanomas either at low frequency or when additional oncogenic mutations are added (e.g., in Pten). We found that as we activate Braf in more melanocytes, such that nevi become more numerous and closely-spaced, the smaller individual nevi become—as if nevi, when close together enough, inhibit each other’s growth. Such behavior is predicted by mathematical models of growth control based on feedback through diffusible signaling molecules. Such models achieve robust control when feedback regulates decisions between self-renewal and progression to alternate cell states or fates. Interestingly, when we look closely at the nevi in this model, we see that there are, in fact, two distinct cell types: highly pigmented nevus body cells and a scattered, lightly pigmented melanocyte population that forms a “veil” around the pigmented cells that had not been observed before. These veil cells are usually not seen unless the melanocyte lineage is fluorescently-labeled with GFP. Single cell RNA-sequencing suggests that these cells likely communicate through ligands and receptors they differentially express.
In this project, Ganesan, Lander and their team seek to understand the role of the nevus body and veil cell types in mouse models that produce both nevi and melanoma,
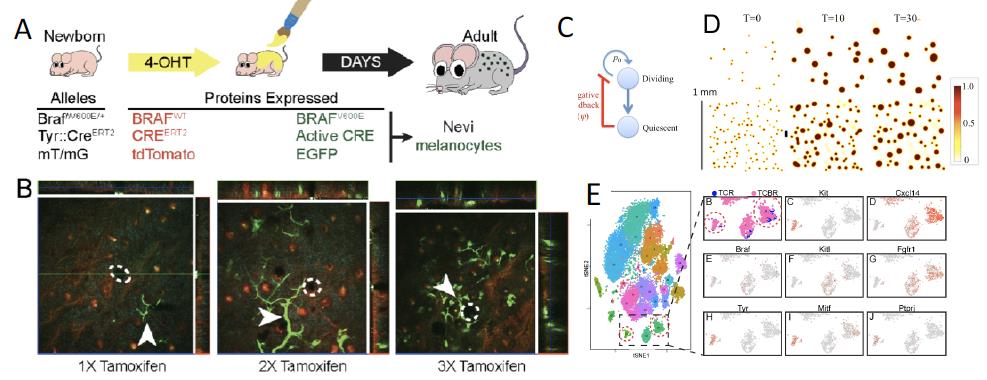
Figure 2. (A) BRAF-mutant (Tyrosinase::CreERT2; BraffloxV600E/+) mice are crossed with ROSAmT/mG mice (TCBR) to GFP-label BRAF-mutant melanocytes. (B) Fluorescence emission (confocal and MPM) 3-D imaging of skin of live mice (as in Fig. 1; total depth = 90, 115 and 190μm for 1x, 2x, and 3x representative stacks, respectively). Green = GFP; red = td Tomato. (C,D). Continuum modeling, in which cells at an arrested (quiescent) lineage stage feedback on the self-renewal probability of dividing cells. Model snapshots (D) show cell density (see heatmap) as a function of time (T= cell cycles) and location. Top and bottom rows are for low and high seeding densities, respectively, that represent different levels of nevi induction. (E) Single Cell RNA-seq of melanocytic nevi. Control mice (Tyrosinase::CreERT2; ROSAmTmG; TCR) and TCBR mice, as labeled. Left: tSNE clustering of gene expression for individual cells. Right: A portion of the tSNE map, showing contributions of TCR and TCBR mice as well as expression of selected marker genes.
and to identify both the nature of how growth is controlled in nevi and the means by which melanoma cells escape from it. The goal of the work is to build a solid molecular and cellular framework on which to base clinical decisions about melanoma prevention, detection and treatment. They are addressing these issues by 1) Explaining feedback growth dynamics in melanocytic nevi; 2) Elucidating how melanomas escape growth control mechanisms that arrest nevi; and 3) Revealing how the immune system targets nevi, and how this affects melanoma development.
This project integrates multiscale mathematical modeling with experiments in mice using a nevus-forming inducible activated Braf model, and a version of the same model that combines Braf activation with inducible loss of one allele of Pten, leading to the reliable production of both nevi and melanoma tumors. We are developing hypotheses that can explain the spatiotemporal dynamics and spatial statistics of nevus and melanoma development in these models, including potential bifurcations that account for the development of both nevi and melanoma in the same mouse. We are investigating the reasons why some cells escape from growth control, while others do not. We anticipate that this is unlikely to be due to a requirement for inactivation of the other Pten allele, and instead believe that escape may more likely be due to the spatial dynamics of collective feedback. The results are expected to shed light on signaling pathways that could be manipulated to prevent or treat melanoma. Live cell imaging, focused laser ablation, immunohistochemistry, and time-course single cell RNA-sequencing are used to identify potential positive and negative feedback regulators that drive the mathematical models, and experiments are used to test model-based predictions concerning the roles that such molecules play. Finally, an investigation of spontaneous regression, which occurs with both mouse and human nevi, provides clues into how the immune system efficiently recognizes melanocyte overgrowth. Since immunotherapy has recently emerged as a promising therapy for melanoma, this study is expected to reveal whether immunotherapy leverages an existing immune program for eliminating nevi, and if so, how that program is carried out, and how melanomas typically escape from it. Such information should aid in developing new prevention and therapeutic strategies for this devastating disease. The integration of this work with Projects 1 and 3 occurs through the use of scRNA-seq as a tool for hypothesis generation and development and through the application of mathematical models that have similar underlying structures (cell state transitions, proliferation and quiescence, positive and negative feedback), but differ in their context (e.g., spatial in Projects 1 and 2 vs. non-spatial in Project 3).